باغبانی نوین
باغبانی و طراحی فضای سبزباغبانی نوین
باغبانی و طراحی فضای سبزدرباره من
 در صورت داشتن نظر، پیشنهاد و انتقاد به ایمیل اینجانب sdoa@chmail.ir اعلام فرمایید.
قبلاً از دقت نظر شما کمال تشکر را دارم.
ادامه...
در صورت داشتن نظر، پیشنهاد و انتقاد به ایمیل اینجانب sdoa@chmail.ir اعلام فرمایید.
قبلاً از دقت نظر شما کمال تشکر را دارم.
ادامه...
گیاهشناسی گردو
گردو درختی است زیبا و سایهافکن که بهعنوان درخت زینتی نیز بهکار میرود. این درخت در مناطق معتدل دنیا رشد و پرورش مییابد. گلهای نر و ماه روی یک پایه قرار دارند. سنبله گلهای نر بهصورت آویخته(شاتون) است ولی گلهای ماده آن بهوضع قائم روی شاخه قرار دارد. میوه گردو، گرد و در درون پردههایی قرار گرفته است. قشری که روی مغز گردو است سبز رنگ و کمی گوشتی میباشد. معمولاً درخت گردو هنگامیکه به سن بیست سالگی میرسد شروع به میوه دادن میکند و بالاترین باروری خود را در سنین پنجاه سالگی دارد. از تمام قسمتهای درخت گردو استفاده طبی بهعمل میآید. درختان گردو درختانی برگریز هستند با برگهای شانهای بطول 90-20 سانتیمتر، تعداد 25-5 برگچه و جوانههای آن دارای درونه حفرهدار هستند.
شاید معروفترین اعضای این رده گردوی سیاه(Juglans nigra) متعلق به شرق آمریکای شمالی و گردوی ایرانی یا معمولی(Juglans regia) که بومی بالکان در جنوب شرقی اروپا، مرکز و جنوب غربی آسیا تا هیمالیا و جنوب غربی چین است، باشد. این دو گونه از نظر چوب قابل توجهی که دارند، دارای اهمیت میباشند. گریبانههای گردو بهعلت دارا بودن مقدار زیادی رنگ زرد- قهوهای، منبع مهمی برای رنگ به شمار میرود. میوههای خشک باغی گردو ارزش بالایی دارد زیرا نگهداری و صادرات آن نیاز به سردخانه و دیگر وسایل پر هزینه ندارد و در ضمن بهعلت خشک بودن هسته آنها صدمه نمیبیند.
گردو در ایران در نقاط غربی و شرقی به فراوانی یافت میشود. گردوی کرمان مرغوبیت بالایی دارد و حتی قدمت آن به 5000 سال میرسد. درختان تنومند خمیده و کهنسال این محصول نشانگر قدمت این درخت میباشد. میزان محصول هر هکتار گردو با بهکارگیری روشهای مکانیزه بهطور متوسط 5/3 تن در هکتار است. انواع رقمهای گردو خود به شش نوع میرسد.
درخت گردو بیشتر به روش پیوند زدن(پیوند اسکنهای، پیوند شکمی یا T معکوس بهدلیل وجود شیرابه) تکثیر میشود. گردو از طریق بذر هم تکثیر میشود ولی میوههایی که بر روی پایههای بذری تولید میشوند استاندارد نبوده و ارزش صادرات ندارد.
دانلود مطلب: ساختار گل و تولید مثل
برای دریافت فایل مطلب در خصوص:
«ساختار گل و تولید مثل»
اینجــــا را کلیک کنید.
ساختار گل و تولید مثل
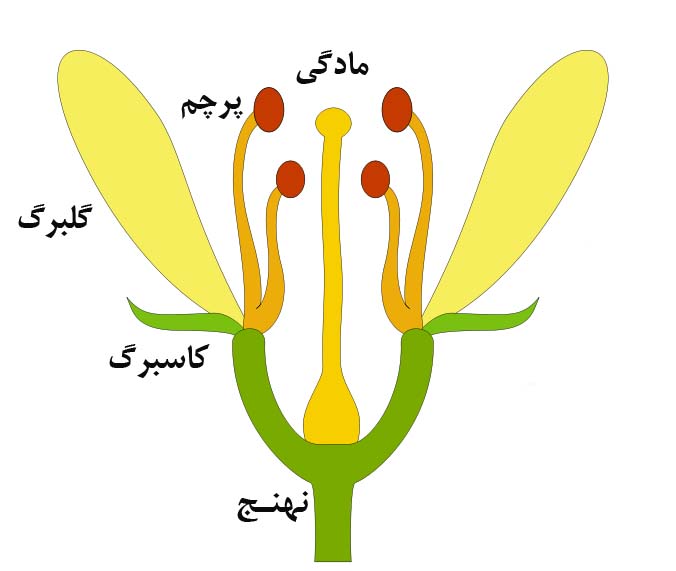

گل ساختارهای زایشی انواع گیاهان نهاندانه است. نهاندانگان گیاهانی هستند که گل و میوه تولید میکنند. گلها معمولاً در هر دو جنس نر و ماده، و رنگ درخشان و جذاب برای جذب حشرات مشاهده میگردد، حشرات با حمل گرده به گلها در تولید مثل جنسی کمک میکنند. تمام گلها رنگارنگ نیستند، این گلها(گلهای فاقد جذابیت و ساده و غیررنگی) معمولاً برای گرده افشانی از باد استفاده میکنند.
اجزای گل:
گل شاخهی تغییر شکل یافتهای است که نهنج معمولاً بخش زیرین آن میباشد. نهنج(B) در انواع گل به رنگهای مختلف دیده میشود. کاسبرگها ساختارهای برگ مانند اطراف گل هستند که عمل محافظت از گل قبل از شکوفایی را برعهده دارند. کاسبرگها(C) معمولاً سبز رنگ هستند. گلبرگ بخش رنگارنگ گل است که میتواند در جذب حشرات و حتی سایر حیوانات کوچک مانند موش، پرندگان و خفاش نقش داشته باشد. رنگ گلبرگ(D) درخشان و جاذب و در گیاهان مختلف متفاوت است. تمام گیاهان گلدار، گل دارند، اما در برخی از آنها رنگ گل جذاب نیست. گلبرگ در این گلها حذف یا کوچک شده است و گیاه برای گرده افشانی متکی بر باد یا آب است.
گل در هر دو جنس مرد و زن بخش تولید مثلی گیاه است. سازه تناسلی زن برچه یا Carpel نامیده میشود. برچه قسمتی از مادگی است که محتوی تخمک می باشد برچهها ممکن است جدا از هم باشند مانند برچههای Ranunculusها و یا پیوسته به هم بوده و از اتحاد آنها تخمدانی چند خانه به وجود آید. در اغلب گلها چندین برچه باهم ترکیب شده به شکل یک مادگی مشاهده میگردد. مادگی(P) در رنگهای مختلف میتواند ایجاد گردد. مادگی دارای سه بخش قابل مشاهده میباشد. کلاله در بالای مادگی قرار دارد که اغلب چسبناک و بر روی آن گرده قرار میگیرد. رنگ کلاله(J) میتواند متفاوت باشد. خامه یا استیل لولهای است که کلاله را به تخمدان متصل میکند. لوله گرده از طریق خامه در پایین این لوله به تخمک میرسد. تخمکها، در تخمدان قرار دارند. گیاهان هر گونه تنها می تواند تخمک همان گونه را بارور نمایند، چون مواد شیمیایی ویژهی هر گونه از باروری تخمکها با گرده انواع دیگر جلوگیری میکند. خامه با(K)، تخمدان با(L) و تخمک با(O) در شکل زیر نمایش داده شدهاند.
مادگی اندام مادهی گل به شمار میآید و ممکن است از یک یا چند برچه تشکیل شده باشد که در حالت اول آن را ساده و در حالت دوم، مرکب مینامند. در مادگی مرکب ممکن است برچهها از هم جدا(آلاله و توتفرنگی) و یا باهم پیوسته باشند مثل زنبق، پامچال، اطلسی.
هر برچه از سه بخش تخمدان، خامه، کلاله تشیکل شدهاست. تخمدان بخش میان تهی است که یک یا چند خانه دارد و تعداد برچههای مادگی مرکب اغلب به تعداد کلالهها و تعداد خانههای تخمدان بستگی دارد. در درون تخمدان ساختارهایی به نام تخمک پدید میآیند. تخمکها حامل گامتهای مادهاند. به نحوه قرار گرفتن تخمک در درون تخمدان، تمکن میگویند. خامه بخش دراز و باریک برچه است که بین کلاله و تخمدان قرار دارد، خامهها ممکن آزاد یا به هم متصل باشند.
در این صورت در وسط ستونی که از اتحاد خامهها ایجاد میشود یک یا چند مجرا به وجود میآید و این مجاری در حقیقت راه عبور لوله گرده برای رسیدن به تخمک است، بخش انتهایی خامه را کلاله میگویند که معمولاً برجسته است و به شکلهای مختلف ظاهر میشود. سطح کلاله اکثرا دارای یاختههای کرک مانند و کوتاهی است که در جذب و نگاهداری گرده موثرند کلاله بعضی از گیاهان مایعی چسبنده و قندی بنام مایع کلاله ترشح میکند. در گیاهانی که گرده افشانی بهوسیله باد انجام میگیرد مانند تیره گندم کلاله منشعب و کرکدار است.
تخمک از سه بخش تشکیل شده است: بافت خورش که تمام یاختههای آن مریستمی هستند، اطراف خورش دو لایه بافت به شکل غلافی بهطرف بالا رشد میکند و نوک خورش را دربرمیگیرد و فقط سوراخ کوچکی در انتها باز میماند که آنرا سفت مینامند. دو لایه بافت پیرامون خورش را پوسته درونی و پوسته بیرونی مینامند.
جفتبندی تخمک و انواع آن:
جفت بندی کناری: در این جفت بندی تخمکها در سطح داخلی تخمدان قرار میگیرند. در تخمدان لوبیا که از یک برچه تشکیل شده، تخمکها در محل اتصال دو لبه برچه قرار دارند و در بنفشه که از سه برچه تشیل شده کنار هر برچه به کنار برچه دیگر متصل میشود و به این ترتیب مادگی سه برچهای تکخانه به وجود میآید و تخمکها در کناره تخمدان در محل اتصال برچهها قرار میگیرند.
جفت بندی محوری: در این جفت بندی کنارههای برچههای تشکیل دهنده مادگی در وسط تخمدان به یکدیگر پیوسته و محور میانی تخمدان را تشکیل میدهند. بنابراین به تعداد برچهها در داخل تخمدان حفره به وجود میآید، و تخمکها به صورت ردیفهایی در طول محور میانی قرار میگیرند، مانند گیاهان تیره سوسن.
جفت بندی مرکزی: این جفت بندی در مادگیهای چند برچه و تکخانه پامچال وجود دارد. تخمکها بر روی ستون آزاد در مرکز تخمدان که از رشد قاعده محل اتصال برچهها حاصل آمده قرار میگیرند، مثل فلفل سبز و انگور فرنگی.
ساختار تناسلی نر:
پرچم اندام نر گیاهان در تولید مثل جنسی میباشد، که از دو قسمت میله و بساک تشکیل شده است. رنگ پرچم(H) متفاوت است. هر پرچم از یک بساک(A) که تولید گرده میکند، میله(F) که بساک را نگهداری می کند. گرده تولید شده بساک توسط حشرات یا حیوانات دیگر به مادگی منتقل میشود.
باروی گیاهان:
تولید مثل جنسی زمانی در گیاهان رخ می دهد که گرده از بساک به کلاله منتقل شود. گیاهان میتوانند خود بارور(خود لقاح) و یا دگر بارور باشند. خود لقاحی زمانی رخ می دهد که گرده از بساک، تخمک همان گل را بارور کند. لقاح متقابل زمانی اتفاق میافتد که گرده به کلاله از یک گیاه کاملاً متفاوت منتقل شده باشد.
هنگامی که تخمک بارور شد، رشد و نمو کرده به دانه تبدیل میشود. گلبرگ پس از لقاح از بین میرود. دیواره تخمدان ممکن است توسعه یافته و قسمت خارجی میوه را تشکیل دهد. انواع بسیاری از میوهها، از جمله سیب و پرتقال و هلو وجود دارد.میوه ساختار محصوری است که از دانه محافظت میکند. میوه در بذرها و لوبیا غلاف مانند هست. هنگامی که شما میوه را میل میکنید، در واقع از تخمدان گل استفاده میکنید.
منبع: http://www.biologycorner.com/worksheets/flower_coloring.html
ترجمه: بهزاد ناقل
انواع ریشه نابجا
1- ریشههای زیرزمینی(Underground Root)
2- ریشههای هوایی(Aerial Root)
2- الف) ریشههای صعود کننده(Climbing roots)
2- ب) ریشههای انگلی(Parasitic roots)
2- ج) ریشههای فتو سنتزکننده(Photosynthetic roots)
2- د) ریشههای نگاهدارنده(Stilt roots)
2- هـ) ریشههای شمعی(Prop or Pillar roots)
تعریف: ریشههای نابجا ریشههایی هستند که منشا آنها ریشهچه جنینی نیست و از قسمتهای دیگر گیاه منشا میگیرند. ریشههای نابجا را میتوان به دو دسته تقسیم نمود. که عبارتند از:
1- ریشههای زیر زمینی(Underground Root): این ریشهها در زیرزمین قرار دارند. ریشه گیاهان تکلپهای و برخی از دولپهایها در این گروه قرار دارند. از تغییرات عمده ریشههای نابجای زیرزمینی میتوان غدهای شدن را نام بردکه در گیاهانی چون مارچوبه Asparagus officinalis و کوکب Dahlia sp. دیده میشود. این ریشه ها را ریشههای غدهای Roots Tuber می نامند.
2- ریشههای هوایی(Aerial Root): این ریشهها بالاتر از سطح خاک و روی اندامهای هوایی(ساقه) تولید میشوند. از مهمترین انواع ریشههای هوایی میتوان انواع زیر را نام برد:
2- الف) ریشههای صعود کننده(Climbing roots): برخی از گیاهان بالا رونده مثل عشقه Hedera helix و پیچ اناری Tecoma sp. به کمک ریشههای ویژه از قیم بالا میروند. این ریشهها معمولاً در بخشهای سطحی قیم فرو میروند و فقط عمل تثبیت گیاه را به عهده دارند. این ریشه به ریشههای صعود کننده موسومند.
2- ب) ریشههای انگلی(Parasitic roots): این ریشهها درگیاهان انگلی مثل دارواش Vischum album، موخور Loranthus sp. و سس Cuscuta sp. وجود دارند. ریشههای انگل در پیکر میزبان فرو میروند وآب، املاح و یا مواد فتوسنتزی را جذب میکنند. این ریشه ها را گاهی مکینه Haustorium نامیدهاند.
2- ج) ریشههای فتوسنتز کننده(Photosynthetic roots): در برخی از ا رکیدههای Orchid اپیفیت ریشههایی وجود دارد که سبز رنگ بوده قادر به انجام عمل فتوسنتز هست. این ریشهها به ریشههای فتوسنتز کننده معروفند.
2- د) ریشههای نگاهدارنده(Stilt roots): در گیاهانی مانند نی Phragmites communis، نیشکرSaccharum officinarum، ذرت Zea mayze و پاندانوس Pandanus ریشههایی وجود دارد که از بخشهای نزدیک به خاک ساقه اصلی خارج شده در خاک فرو میرود و در قائم ماندن ساقه آن را کمک مینماید. این ریشهها ریشههای نگاهدارنده نام دارند.
2- هـ) ریشههای شمعی(Prop or Pillar roots): این ریشه ها در لور(مکر زن) Ficus bengalensis و انجیر معابد Ficus religusa وجود دارد. در این گیاهان از شاخه های درخت ریشههایی تولید می شود که در زمین فرو میروند و از شکسته شدن شاخهها محافظت میکنند. این ریشهها بعداً ضخیم شده و شبیه به تنه میشوند. بدین ترتیب به نظر میرسد که چنین درختانی دارای چندین تنه هستند. این ریشه را ریشههای شمعی می نامند.
هورمونهای گیاهی
واژه هورمون به مواد معینی اطلاق میشود که در بخشی از موجود زنده ساخته شده و پس از انتقال اثرات فیزیولوژیکی محسوسی در دیگر قسمتهای آن به جا میگذارد و در تراکمهای بسیار کم فعالند. این تصور کلی در اصل در قلمرو فیزیولوژیکی حیوانی به وجود آمده و این واژه هورمون از ریشه یونانی به معنی تهیج کردن گرفته شده است. در گیاهان ترکیبات مترادف ولی از نظر شیمیایی کاملاً متفاوت یافت میشود و واژه هورمون به طور صحیح آنها را در بر میگیرد. هورمونهای گیاهی که اغلب فیتوهورمون خوانده میشود در بافتهای مریستمی و یا لااقل جوان از هر نوع ساخته میشوند و غالباً اثرخود را پس از انتقال میگذارند که تا حدودی دورتر از بافتی که ساخته شدهاند، میباشد. هورمونها با آنزیمها و ویتامینها و DNA در این خاصیت مشترکند که به غلظت بسیار کم یا ناچیز باعث ایجاد اثرات فیزیولوژیکی عمیق میشوند. اصولاً واژه هورمون باید به ترکیباتی محدود شود که به طور طبیعی در درون موجود زنده ساخته میشود لذا در تعریف هورمون گیاهی میتوان گفت مواد آلی میباشد که مواد غذایی نبوده و توسط گیاهان تولید میشود و در غلظتهای کم فرآیند فیزیولوژیکی را تنظیم میکند.
آنها در درون گیاه، از محل تولید به محل اثر، انتقال مییابد اما گاهاً موادی که معلوم نیست در گیاه وجود داشته باشد اثرات مشابه و بعضی اوقات عیناً نظیر یکی از هورمونهای طبیعی گیاهی را دارندکه از نام نهادن هورمون گیاهی میبایست خودداری نمود که البته واژه مناسب برای این چنین ترکیباتی که اثر هورمون مانند روی گیاه دارند تنظیم کننده رشد میباشد و در تعریف آن میتوان گفت ترکیبات سنتز شده یا هورمونهای گیاهی هستند که فرآیندهای فیزیولوژیکی را تغییر میدهد. این مواد تقلید کننده از هورمونها، تاثیر روی سنتز هورمونها و از بین بردن و یا انتقال و یا(به احتمال) تغییر دادن محل تاثیر هورمونی رشد را تنظیم میکند. با این وصف برای متمایز کردن آنها میتوان گفت تمام هورمونها، تنظیم کننده رشد هستند اما تمام تنظیم کنندههای رشد هورمون نیستند.
در حال حاضر در دنیا پنج گروه مختلف هورمونهای گیاهی شناخته شده که بسیاری از آنها دارای کاربردهای عملی متعدد و مهمی در کشاورزی بویژه باغبانی هستند این گروهها عبارتند از: اکسینها، جیبرلینها، سایتوکنینها، اتیلن، و مواد بازدارنده رشد.
فلاونوئیدها
واژه فلاونوئید به مفهوم وسیع تمام رنگدانههای گیاهی را در بر میگیرد. نام فلاون از کلمه لاتین فلاووس به معنای زرد گرفته شده است. در گیاهان عالی به ویژه نهاندانگان حدود 4000 نوع فلاونوئید شناسایی شده که بیش از 90 نوع فلاونوئید در مرکبات و بیش از 30 نوع در خانواده کمپوزیته موجود است. تاکنون هیچ نوع فلاونوئید در جلبکها گزارش نشده است. در میان ترکیبات فنولی فلاونوئیدها کلیه اعمال متابولیتهای ثانویه را درگیاهان دارا میباشند. فلاونوئیدها از مشتقات فنیل پروپانوئید هستند که دارای ساختمان 15 کربنه میباشند. در بیشتر فلاونوئیدها حلقه A متادیهیدروکسیل یا متاتریهیدروکسیل میباشد در مقابل حلقه B دارای یک یا دو یا سه عامل هیدروکسیلی میباشد. این اختلاف ناشی از مبدا بیوسنتز دو حلقه میباشد. حلقه A از سه مولکول اسید استیک متراکم و حلقه B از قندها در مسیر شیکمیک مشتق شده است.
خواص درمانی فلاونوئیدها:
فلاونوئیدها باعث ایجاد رنگ در گلها، میوهها و گاه برگها میشوند. همچنین به علت خاصیت جذب حشرات در گردهافشانی و باروری گیاهان نیز موثر میباشند. فلاونوئیدها باعث افزایش مقاومت به عوامل بیماریزا در گیاهان میشوند همچنین جذب کننده قوی اشعه ماورای بنفش(340-250 نانومتر) میبا شند. در ساختمان اندامهای جنسی و رویشی دانه گرده از طریق اثر بر عمل ژنها و آنزیمها اثر مثبت دارند. این ماد همچنین باعث شلاته کردن برخی یونهای فلزی مانند آهن ومس میشوند. فلاونوئیدها از طریق مهار عناصر کاتالیزشده از اکسیداسیون جلوگیری میکنند. از دیگر خواص این مواد میتوان به خاصیت جمع کنندگی مواد زاید اشاره کرد.
فلاونوئیدها مهار کننده سلولهای سرطانی از طریق بیان ژن، افزایش دهنده ایمنی بدن(آنتی اکسیدان)، ضد ویروس، ضد باکتری، ضد تورم، ضد التهاب، ضد آلرژی، ضد موتاسیون، کاهش دهنده نفوذپذیری و شکنندگی مویرگها میباشند. همچنین در کشور چین برای درمان هپاتیت از فلاونوئیدها استفاده میشود. 45 درصد فلاونوئیدهای مرکبات بهطور ویژه جذب ویتامین C را بهبود میبخشند. افراد سیگاری و آنانی که تحت فشار استرس هستند، به میزان زیادی از ویتامین C و فلاونوئیدهای مرکبات نیاز دارند. فلاونوئیدهای مرکبات میتوانند به فرم ترکیباتی جهت کنترل افراد سیگاری یا به شکل آمپولهایی برای ورزشکاران مورد استفاده قرار گیرند.
معرفی تعدادی از گیاهان حاوی فلاونوئید:
بومادران، سرخ ولیک، جینکو، آویشن، گل راعی، بابونه رومی، انار، چای، زالزالک، برگ بو، گل ساعتی، مرکبات، شیرین بیان و ... .
علت تغییر رنگ برگ ها
دست هنرنمای طبیعت در فصل پاییز آشوبی از رنگ های قرمز و زرد در دل جنگل بر پا می کند. درک این تصاویر زیبا، بی قوه تخیلی و چالاکی نیز میسر است و هر کس به سادگی قادر به ادراک آن می باشد. هــر سال با آغاز فصل پاییز با دیدن درختان از سرور مبهمی سرشـار می شویم. در حالی که می دانیم این لذت، زودگذر و بی ثبات خواهد بود. در مدت کوتاهی برگ ها به آرامی از ماوای تابستانی خود جدا شده و زمین جنگل را مفروش می کنند. بسیاری از مردم گمان می کنند که یخبندان باعث تغییر رنگ برگ ها می شود. در صورتی که این تصور درست نیست زیرا رنگ بعضی از برگ ها قبل از فصل انجماد شروع به تغییر می کند. ذهن خیال برخی از مردم هند علت آن را شکار توسط شکارچیان آسمانی(دب اکبر) می پندارند، بدین معنا که خون شکار بر روی درختان جنگل چکیده و بسیاری از برگ ها را به رنگ قرمز درآورده است و برگ های درختان دیگر نیز بر اثر چربیی که به هنگام پخت گوشت آن توسط شکارچیان بر روی درختان پاشیده شده به رنگ زرد درآمده اند ولی حقیقت امر این است که فرآیندهای شیمیایی که به هنگام آماده شدن گیاه برای فصول زمستان در آن به وقوع می پیوندند تغییر رنگ برگ ها را باعث می گردند. برگ ها در طول فصل بهار و تابستان مانند کارخانه هایی عمل می کنند چرا که تهیه غذای لازم برای رشد درختان را به عهده دارند. ساخت مواد غذایی توسط سلول های ریز بی شمار برگ، که متعلق به بافت سننتزی مزوفیل که میان دو بشره زبرین و زیرین برگ است، و بعضی اوقات این بافت کلرانشیم نامیده می شود صورت می گیرد، زیرا سلول های این بافت دارای کلروپلاست های بی شمار است که این کلروپلاست ها حاوی اجسام سبز کوچکی می باشند که کلروفیل نام دارند و رنگ برگ را موجب می گردند و در میان سلول های فتوسنتزی برگ رگبرگ های بی شماری قرار گرفته که از سلول های آوند چوبی و آبکش تشکیل یافته است که آب و مواد معدنی را به درون برگ آورده، فرآورده های فتوسنتزی را به خارج از آن هدایت می کند. رگبرگ های مزبور نهایتاً به رگبرگ اصلی متصل می شوند که پس از عبور از دمبرگ به سیستم آوندی اصلی گیاه می پیوندند کلروفیل ها غذا را برای گیاه از طریق ترکیب نمودن کربن به دست آمده از هوا، با ئیدروژن، اکسیژن و مواد معدنی گوناگون موجود در آب و طی فرآیند فتوسنتز یا نور ساخت فراهم می کنند. در پاییز هنگامی که هوای سرد موجب کاهش فرآیند های حیاتی می گردد فعالیت برگ به پایان می رسد، به بیان ساده تر اینکه اجزاء دستگاه کارخانه برگ پیاده می شوند، کلروفیل به مواد متنوعی که از آنها ترکیب یافته تجزیه می شود و مواد غذایی قابل دسترس به منظور ذخیره سازی برای استفاده در بهار به تنه درخت منتقل می گردد و آنچه که در حفره های سلولی برگ ها باقی می ماند نوعی ماده آبکی است که در آن مقداری قطرات چربی، بلور و تعداد معدودی اجسام زرد رنگ که به شدت نور را منکسر می سازند مشاهده می شوند که عامل زرد شدن رنگ برگ ها در پاییز به شمار می روند.
غالباً میزان قند در برگ ها بیش از آن است که بتواند به سرعت به درون درخت انتقال یابد. در این حالت ترکیب شیمیایی آن با سایر مواد، رنگ های متنوعی را به وجود می آورد که بین قرمز روشن و قرمز مایل به قهوه أی در نوسان است. در مخروط داران که برگ های خود را در پاییـز از دست نمی دهند به قهوه ای روشن بدل می شود که خود عامل روشن تر شدن رنگ در بهار می باشد. در مدت زمانی که برگ ها تغییر رنگ می دهند بر اثر عواملی از محیط که عبارتند از کوتاه شدن طول روز و کاهش دما، افتادن برگ از درختان خزان پذیر آغاز می گردد. در نقطه ای که دمبرگ به تنه چسبیده لایه ای از سلول های خاص تکامل می یابند و به تدریج بافت هایی که برگ را نگه می دارند سست شده و برگ ها جدا می شوند. سلول های پارانشیمی لایه ریزش؛ اغلب از سلول های اطراف کوچکتر است، حتی عناصر آوندی کوتاه بوده در دسته های آوندی در ناحیه ریزش فیبر وجود ندارد. این ویژگی های تشریحی ناحیه ریزش را به نقطه ضعیفی مبدل می سازد. قبل از افتادن برگ تغییرات بی شماری در ناحیه ریزش روی می دهد. تقسیم سلولی غالباً انجام می گیرد و لایه أی از سلول هایی به شکل آجر در سراسر عرض پایه دمبرگ به وجود می آید. تغییرات فعال متابولیسمی در سلول های ناحیه ریزش، سبب تجزیه جزیی دیواره سلول یا تیغه میانی می شود و سلول ها بدین ترتیب از هم جدا می گردند و سنگینی برگ و یا وزش باد در نهایت اتصال آوندی را گسسته، برگ از درخت می افتد. هم زمان با جدا شدن برگ ها طبیعت جای بریدگی را ترمیـم می نماید، به این شکل که وقتی برگ بر اثر وزش باد یا وزن خود فرو می افتد یک لایه چوب پنبه أی روی بقایای دمبرگ تشکیل می شود که گیاه را از حمله میکروبی محافظت می کند و اتلاف آب را محدود می سازد. آوندهای چوبی بر اثر نفوذ سلول های پارانشیمی مجاور یعنی نفوذ تیل به دورن آنها، مسدود شده عمل انسداد کامل می گردد و محلی در ساقه که برگ روی آن رشد کرده توسط یک زخم مشخص می شود. ریزش برگ پیش بینی خردمندانه دیگر طبیعت برای فصل زمستان می باشد. درختان پهن برگ برگ های خود را از دست می دهند و در نتیجه شاخه های آنها با سهولت بیشتری سنگینی برف و یخ زمستان را تحمل می کنند. در نقاطی که به ندرت برف و یخ به چشم می خورد برخی از پهن برگان به طور دائم سبز می باشند.
مخروطیان(کاج ها، سروها و نرادها) زمان مشخص برای ریز برگ ندارند. برگ های آنها سوزنی یا فلسی شکل می باشد( فرم آنها با ریزش برف مطابقت دارد). طبیعت سیستم کارآمدی برای دوباره به چرخه درآوردن شاخ و برگ های فروریخته دارد و از طرف دیگر مقدمات یک خاک جنگلی حاصلخیز را فراهم می آورد. گرچه مواد غذایی آماده شده در حفره های سلولی برگ ها در فصل پاییز به داخل درخت بازگردانده می شوند، اما مواد معدنی که در طول فصل تابستان درون دیواره های سلولها تزریق شده اند همچنان باقی می مانند. بنابراین برگ های ریخته شده حاوی مقادیر نسبتا زیادی عناصر ارزشمند مانند نیتروژن و فسفر که در واقع بخشی از خاک به حساب می آیند می باشند. تجزیه برگ ها لایه های بالایی خاک را از طریق بازگردان عناصری که توسط گیاه قرض گرفته شده غنی گردانده و نه تنها موجب تغذیه درختان درختچه ها و بوته ها در طول سال می گردد بلکه آنها را در مقابل مشکلات دمایی فصل تابستان و زمستان محافظت می کند و از طرف دیگر موجب تثبیت خاک در محل می گردد و به طور همزمان مقدمات تجمع آب را فراهم می سازد. اگر مشتی از این خاک تیره رنگ را بررسی کنید مشاهده خواهید کرد که حتی در یک دوره خشکسالی مرطوب تر و نرمتر از خاک های زمین های اطراف می باشد(هوموس جاذب الرطوبه) به هر حال اگر اجازه داده شود که آتش در داخل جنگل پیشروی نماید و برگ ها بسوزند ارزشمندترین عناصر تقویت کننده بر اثر حرارت تبدیل به گاز می شوند و به هوا می روند و هوموس با ارزش تخریب می گردد. جنگل هایی که سطح شان به طور مداوم سوزانده می شود به زودی خاک حاصلخیز خود و توانایی جذب آب و نگهداری رطوبت را از دست خواهند داد حتی اگر هیچ صدمه چشمگیری به درختان استوار وارد نشود.
منبع: سایت های اینترنتی
گیاه شناسی ساقه
ساقه گیاهان دارای رشد طولی و رشد قطری است. تمام ساقهها رشد طولی دارند ولی رشد قطری در همه آنها دیده نمیشود. هر دو نوع رشد نتیجه تقسیم یاختههای مریستمی و تولید یاختههای جدید است. رشد طولی مقدم بر رشد قطری است و در ساقههایی که دارای هر دو نوع رشدند ابتدا رشد طولی را آغاز میکنند. به همین دلیل رشد طولی و رشد نخستین و رشد قطری را رشد پسین مینامند.
پیدایش بافت های نخستین ساقه:
در مریستم ساقه سه نوع بافت مریستم نخستین به نام های پروتودرم، مریستم زمینه، و پروکامبیوم تشخیص داده میشود. این سه نوع بافت چند میلیمتر پایینتر از مریستم انتهایی قرار دارند و مستقیما از تمایز یاختههای مریستم انتهایی به وجود میآیند. از تمایز این سه نوع به ترتیب بافت های زیر پدید میآیند:
پروتودرم= بشره را تولید میکند.
مریستم زمینه= بافت های نخستین پوست، مغز، و اشعه مغزی را تولید میکند.
پروکامبیوم= ابتدا بافت های چوبی و آبکشی نخستین را به وجود میآورد و بعدها کامبیوم آوندی را تولید میکند. پروکامبیوم از خارج آوندهای آبکشی و از داخل آوندهای چوبی را ایجاد میکند.
ساختار بافت های نخستین دو لپهای ها و بازدانگان:
بشره، پوست، آندودرم، استوانه مرکزی، مغز و اشعه مغزی
بشره شامل یک ردیف یاختههای بدون کلروپلاست و نشاستهای ناحیه استوانهای پوست است و معمولاً از بافت پارانشیم به وجود آمده است. آندودرم از یک لایه یاخته تشکیل شده است که استوانهای تو خالی را تشکیل میدهد در یاختههای معمولی آندودرم نوارهای محتوی چوب و چوب پنبه به صورت نوار کاسپار دیده میشود. آندودرم ساقه چندان واضح دیده نمیشود. استوانه مرکزی به صورت استوانهای پوسته یا گسسته درون پوست قرار دارد. در استوانه مرکزی بافت آبکشی و بافت چوبی وجود دارد که بافت آبکشی معمولاً در خارج قرار میگیرد و شامل یاختههای آبکشی، یاختههای همراه، فیبر و پارانشیم است و دیگری بافت چوبی که در داخل قرار دارد شامل یاختههای چوبی، تراکئید، فیبر و پارانشیم است. در حالتی که استوانه مرکزی یکپارچه نیست، هر نوار یک دسته آوندی خوانده میشود بین آوندهای آبکش و آوندهای چوبی ساقه دو لپهای های چوبی و بازدانگان بافت کامبیوم آوندی در یک لایه قرار دارد. بخش بیرونی استوانه مرکزی را دایره محیطی تشکیل میدهد. مغز بخشی کم و بیش استوانهای است که در مرکز ساقه قرار دارد و از بافت پارانشیم تشکیل شده و انشعابات آن بنام اشعه مغزی فواصل بین آوندها را پر میکند.
ساختار نخستین تک لپهای ها:
در ساقه علفی تک لپهای ها بافت های چوبی و آبکشی به صورت دستجات آوندی پراکندهاند و تراکم آنها در نزدیکی های پیرامون ساقه بیشتر است. بافت چوبی اغلب در سمت درونی ساقه و در زیر بافت آبکشی مربوط قرار دارد. آوندهای چوبی و آبکشی توسط بافتی بنام غلاف آوندی احاطه شدهاند. مغز اکثراً تحلیل رفته است به علت عدم رشد پسین کامبیوم ندارند و در بافت هایی مانند نخل ساقه کلفت میشود اما چوبی نیست.
پیدایش بافت های پسین در ساقه:
رشد پسین در اثر فعالیت کامبیوم چوب آبکش صورت میگیرد این لایه بافت آبکشی پسین را به طرف خارج و بافت چوبی را پسین را به طرف داخل تولید میکند. افزایش قطری ساقه سبب ایجاد شکاف هایی در بشره و پوست میگردد در این صورت نقش های حفاظتی و ذخیرهای بشره و پوست به وسیله بافت های چوب پنبه و فلودرم انجام میگیرد. این بافت ها از کامبیوم فلوژن حاصل میشود.
ساختار درونی پسین ساقه دو لپهای های چوبی و بازدانگان:
کامبیوم چوب آبکش به طرف خارج آبکش پسین را تولید میکند و به طرف داخل بافت چوبی پسین را تولید میکند تناوب فعالیت کامبیوم چوب آبکش سبب ایجاد چوب های بهاره و تابستانه میشود. که در مجموع حلقه سالانه را به وجود میآورند سن درخت را میتوان با شمارش حلقه در سطح مقطع تنه درخت تعیین کرد. با افزایش سن درخت ویژگی های چوب قسمت مرکزی ساقه تغییر میکند و نقش هدایت شیره خام را از دست میدهد این چوب سختتر و تیرهتر از چوب پیرامون است و آن را قلب چوب مینامند. چوب پیرامونی قلب چوب تغییر نمیکند و همچنان هدایت شیره خام را بر عهده دارد و چوب پیرامونی را چوب شیره بر مینامند.
به لحاظ استقرار جوانه به روی ساقه شامل انواع: جوانه انتهایی، جانبی، فرعی و نابجا می باشد و به لحاظ اندامی که تولید میکنند شامل جوانه گل، برگ، و مرکب است.
ساقههای چوبی: در ساقههای چوبی مانند گردو در نوک شاخه جوانهای به نام جوانه انتهایی و در طول شاخه آن جوانههای جانبی وجود دارد و در پایین هر جوانه اثر آوند و اثر برگ دیده میشود. در طول ساقههای چوبی در محل ارتباط بافت های آوندی برگ و جوانه گره وجود دارد و در سطح ساقه برآمدگی های کوچکی به نام عدسک دیده میشود.
جوانه از یاختههای مرسیتمی تشکیل شده است. این یاختهها در برابر عوامل نامساعد محیط بسیار حساساند و نیاز به محافظت دارند به همین مناسب اغلب جوانهها از برگ های تغییر شکل یافتهای به نام پولک تشکیل شدهاند. جوانه های گیاهان علفی و معدودی از گیاهان چوبی پولک های حفاظتی ندارند و آنها را جوانه برهنه می نامند.
ساقه گیاهان دو لپهای علفی: ساختار ظاهری این گیاهان شبیه به ساقه جوان گیاهان چوبی است. اما جوانهها برهنه و در سراسر عمر گیاه فعالاند. برگ های این گیاهان نمیریزند و در نتیجه اثر برگ ها و اثر بافت های آنها روی ساقه دیده نمیشود.
ساقه گیاهان تک لپهای: ذرت و نخل دو نمونه از گیاهان تک لپهای هستند. ذرت تک لپهای علفی است که ساقه آن از نیام برگ ها پوشیده شده است. اگر نیام را جدا کنیم در ساقه آن گره و میانگره دیده میشود ساقه در محل گرهها کمابیش تخم مرغی شکل و در یک نقطه فرورفته است.
ساقه نخل ها: دارای جوانه انتهایی مخروطی شکل بسیار بزرگند. که برگ های جدید و گل از آن تولید میشوند. اگر نقطه رشد انتهایی آسیب ببیند گیاه میمیرد. برگ ها نزدیک به هم در بالای ساقه تولید میشوند در نتیجه میانگرهها کوتاهند. در ساقه نخل جوانه، گره و میانگره به وضوح دیده نمیشوند. ساقه نخل رشد قطری ندارد و قطر آن از بالا به پایین یکسان است. علت قطور بودن ساقه نخل بزرگ شدن یاختهای پارانشیمی ساقه و تمرکز ماده چوب و سایر مواد دیگر در دیوارههای آنهاست.
ساقههای تغییر شکل یافته: تغییر شکل ساقه اغلب با تغییر نقش آن همراه است. در هر حال ساقه با هر شکل و نقشی ویژگی های ساختاری خود را داراست. یعنی گره، میانگره و بافت های مشخصی دارد. مهمترین ساقههای تغیر شکل یافته عبارتند از:
ساقه هوایی خزنده: این ساقهها عموما در سطح زمین به طور افقی رشد میکنند و دارای میانگره بلندند(توت فرنگی( برگ ها ریز و پولک مانند و برگ و گل در گرههای معین یا در محل گرههای که با زمین تماس حاصل میکنند تولید میشود.
ساقه زیرزمینی: این ساقهها اندام های ذخیرهای گیاه به شمار میروند. با استفاده از مواد ذخیرهای که در طی سال اول در آنها جمع میشود ساقه هوایی جدیدی در سال بعد رشد میکند. ساقههای زیرزمینی به شکل ریزوم، غده پیاز(سوخ) دیده میشود. در ریزوم ساقهها استوانه ای شکلاند و در زیر زمین به طور افقی رشد میکنند این ساقهها باریک و گوشتی و دارای اندوخته غذایی هستند. ساقهها دارای گره، میانگره، برگ های متعددند. جوانهها در پایه برگ های پولکی اندام های هوایی را تولید میکنند و در گونههای زنبق انتهای در حال رشد ریزوم برگ و گل تولید می کند. ریشه در محل گره ها تولید می شود.
در ساقه های غده ای انتهای متورم ریزوم را غده مینامند. سیب زمینی یک غده است. بوته سیب زمینی سه نوع ساقه دارد: ساقههای هوایی معمولی، ریزوم باریک و انتهای متورم آن همان غده است. غده سیب زمینی
دارای گره، میانگره، جوانه جانبی و یک جوانه انتهایی است. گروهی از جوانهها یک چشم را تشکیل میدهند. چشم های بر روی غده به وضع مارپیچی قرار دارند. هر چشم موقعیت یک گره را نشان میدهد و در ساقه پیازی ساقهها کوتاه و ضخیماند، به طور افقی رشد میکنند و غذای اندوخته یا در ساقه کوتاه مثل گلایول و سیکلامن یا در پولک های برگ مانند اطراف آن مثل نرگس جا دارند.
ساقه پیچنده یا پیچک ها: ساقه پیچنده دراز و باریک است و بافت استحکامی دارد. در تماس با هر یک تکیه گاه به دور آن میپیچند مانند پیچک انگور، نیلوفر، چسبک، پیچک ها؛ در بخش انتهایی خود رشد سریع دارند.
ساقه برگ نما: به شکل ظاهری ساقه برگ نما همانند برگ است. این ساقهها سبز رنگند و نقش برگ را هم انجام میدهند و سطح این ساقهها ممکن است گل، میوه و برگ به طور موقت ظاهر شود. مانند کوله خاس، مارچوبه.
ساقه گوشتی: در عده ای از گیاهان فرایند ساختن غذا محدود به ساقه میشود، زیرا برگ ها بسیار تحلیل رفتهاند. این گیاهان در مواقع بارندگی مقدار قابل ملاحظهای آب را در ساقه گوشتدار خود ذخیره میکنند و در فصل های بیآبی از آن استفاده میکنند. ساقه گیاهان کاکتوس، فرفیون، علف شیر از جملهاند.
ساقه خار نما: اغلب خارهای گیاهان، ساقه تغییر شکل یافته یا زایده ساقهاند. اما خارهایی که از تغییر شکل برگ ها حاصل شدهاند در بعضی گیاهان مانند زرشک و اقاقیا دیده میشوند. نمونه ساقه خار نما در گیاه خار مصری و لالیک دیده میشوند. سطح خارها دارای برگ است که دلیلی است بر ساقه بودن خار.
منبع: سایت های اینترنتی
قوانین نامگذاری بینالمللی گیاهان
در قرن هیجدهم با افزایش اسامی گیاهان، گیاهشناسان اروپایی بر آن شدند که قوانین را جهت نامگذاری علمی گیاهان به وجود آورند. اولین قدم توسط لینه در سال 1751 برداشته شد و در اواخر قرن هیجدهم با حفظ حق تقدم Priority برای اسامی قدیمی گیاهان فعالیت های بعدی شروع شد. به این ترتیب گیاهشناسان به تدریج سیستم نامگذاری را به وجودآوردند که گیاهان به گروه های مشابه تقسیم شوند و برای نامگذاری آنها از اسامی لاتین استفاده شود و یا هر اسمی که از زبان های دیگری گرفته شده باشد، به صورت لاتین برگردانده شود. علت اینکه زبان لاتین به عنوان زبان علمی و رسمی برای نامگذاری گیاهان به کار برده شد، به این جهت بود که اولاً از زمان های قبل از لینه و همچنین در عصر لینه لاتین زبان علمی دانشمندان بود و از طرفی چون زبان لاتین در مطالعه و مکالمه مورد استفاده نمیباشد مانند زبان های دیگر کمتر دچار تغییر و تحول میگردد. از طرف دیگر به کار بردن اسامی عامیانه در زبان های مختلف، بینالمللی نبوده، در نتیجه در بسیاری نقاط دنیا ممکن بود گیاهی (گونهای) دارای اسامی متعدد باشد و یا اسامی عامیانه و محلی، بدون تمایز بر روی جنس و گونه گذاشته شود. مثلاً سروکوهی Juniperus در بعضی کتاب های فارسی به اسامی عرعر، ارس، کوکلان و آردج ذکر شده است و یا درخت سرو Cupressus به نام های متنوع سرو، ورس، اورس، درخت آزاد و سرو آزاد ذکر شده است و یا درخت لیلکی Gleditschia در شمال ایران دارای اسامی مختلف به نام های لیلک، لالکی، لک و کرات میباشد و به همین طریق در زبان های دیگر. با توجه به مسائل فوق، سیستم نامگذاری به وجود آمد که به کار بردن اسامی علمی را به صورت استاندارد شدهای در عالم گیاهی و جانوری متداول کرد.
در نتیجه گیاهشناسان و جانورشناسان امروزی نیز در تمام کشورها قوانین بین المللی خاصی را برای نامگذاری علمی گیاهان و جانوران به کار میبرند که به نام قوانین نامگذاری بینالمللی گیاهشناسی International code of Botanical Nomenclature برای گیاهان ICNB و قوانین نامگذاری بین المللی جانورشناسی International code of Zoological Nomenclature برای جانوران ICNB میباشد. هدف از قوانین بین المللی نامگذاری گیاهان، ایجاد روش نامگذاری ثابت برای گروه های تاکزونومیکی میباشد که از به کار بردن اسامی و نام هایی که باعث اشتباه و یا ابهام و یا اغتشاش در علم میشود، جلوگیری نماید. اولین بار کنگره گیاهشناسی بینالمللی توسط گیاهشناس معروف سویسی آلفونس دوکاندول درسال 1867 در پاریس تشکیل شد که در این کنگره قوانین زیر به عنوان زیربنای نامگذاری گیاهان پایه ریزی شد:
هیچ گیاهی نباید بیش از یک اسم علمی داشته باشد.
نام گونه باید همراه با نام جنس ذکر گردد.
برای هر اسم باید شرحی نوشته شود و یا وسیله دیگری جهت شناسایی گیاه مزبور فراهم گردد و ذکر تاریخ حتمی است.
دوگونه گیاهی نباید دارای یک اسم علمی مشابه باشد.
نام گیاه باید همراه با نام مؤلف آن گیاه باشد.
با توجه به تغییراتی که به تدریج در قوانین نامگذاری لازم به نظر میرسید، کنگرههای بعدی در سنوات 1910،1907،1905،1892 تشکیل گردید و مهمترین کنگره در سال 1905 در وین بود که مسائل مهم مربوط به نامگذاری را مورد بحث قرار داد. از آن جمله قرار شد برای کلیه اسم های جدید از اول ژانویه 1908 شرحی به لاتین نوشته شود و به تدریج در طول تاریخ گیاهشناسی قوانین کلی و بسیار مهمی برای نامگذاری گیاهان در نظر گرفته شد که تمام گیاهشناسان دنیا بایستی این مقررات و قوانین بینالمللی را رعایت نمایند. شرح کامل قوانین و کد نامگذاری توسط لورنس Lawrence (1951)، اسمیت(1957)، استافلو Stafleu (1968)، یا اصطلاحات مورد استفاده توسط استرن Stearn در کتاب واژهنامه گیاهشناسی لاتین Botanical latin در سال 1973 به طور کامل ارایه شده است.
امروزه گیاهشناسان دنیا کد بینالمللی نامگذاری گیاهان را به کار میبرند که به زبان های انگلیسی، فرانسه و آلمانی نوشته شده است. این کد شامل چند اصل مهم و مشخص میباشد که عبارتند از:
اصل اول: اسامی علمی گیاهشناسی Botanical Nomenclature مستقل از اسامی علمی جانورشناسی Zoological Nomenclature میباشد و ممکن است اسامی و نام هایی که برای گیاه به کار برده میشود در مورد جانور نیز همان اسم به کار برده شود. به عنوان مثال اسم کوکروپیا Cocropia هم برای جنس حشره شب پرک بید و هم برای یک جنس بزرگ از گیاهان گرمسیری از خانواده توت Moraceae به کار برده میشود. کوریدالیس Corydalis جنس گیاهی از خانواده شاه تره Fumariaceae و جنس حشره مورچهخوار کوریدالیس از حشرات زیر خانواده کوریدالیدا Corydalidae راسته بال توری مگالوپترا Megaloptera میباشد.
اصل دوم: اسامی درخواستی برای گروه های تاکزونومیکی براساس نمونه تیپ Type specimen تعیین شده است. اصل مربوط به تیپ این است که اسم هرگونه بایستی روی نمونه به خصوص و مشخصی گذارده شده باشد که به نام نمونه تیپ نامیده میشود. نمونه تیپ هر جنس یک گونه و تیپ هر خانواده یک جنس میباشد و غیره.
اصل سوم: اسم علمی یک گروه تاکزونومیکی بر اولویت انتشار اسم آن گروه بنا شده است یعنی اسم صحیح اسمی است که طبق قوانین و کد بینالمللی زودتر چاپ شده باشد. به همین منظور اولویت یا حق تقدم در گیاهشناسی از اول مه 1753 برای اسامی گیاهان آوندی با انتشار «گونههای گیاهان Species plantarum» لینه آغاز شد.
اصل چهارم: هر گروه تاکزونومیکی با موقعیت خاص خود در هر طبقه فقط میتواند دارای یک اسم باشد که طبق قوانین نامگذاری قدیمیترین اسم خواهد بود(به جز در مورد گونه).
اصل پنجم: اسامی علمی گروه های تاکزونومیکی بایستی به صورت لاتین نوشته شده باشد که شامل اسامی جنس، گونه و سایر گروه ها میباشد.
اصل ششم: قوانین مربوط به نامگذاری عطف به ماسبق میشود، مگر آن که صریحاً محدود شده باشد.
اصولی که پایه سیستم نامگذاری گیاهان را تشکیل میدهد شامل قوانین Rules و توصیهها یا پیشنهادات Recommendations میباشد. منظور از قوانین این است که اصطلاحات علمی گذشته در دستور مشخصی گذاشته شود که پایهریزی برای آینده باشد و هدف از توصیهها بیشتر روشن نمودن مسائل و یک شکل نمودن اسامی علمی گیاهان میباشد.
منبع: کتاب اصول و روش های رده بنـدی گیاهـان دانشگاه پیام نور
نیاز نوری گیاهان
همه گیاهان به یک اندازه به نور نیازمند نیستند. بعضی از گیاهان نیاز به نور فراوانی دارند و بعضی دیگر به نور کمتری نیاز دارند. بنابراین گیاهان را از نظر نیاز نوری به سه گروه بزرگ تقسیم میکنیم:
گیاهان روز بلند LDP: Long Day Plants برای به گل رفتن بین 10 تا 14 ساعت به نور نیاز دارند، مانند گیاهان فصلی تابستانه نظیر آحار، اطلسی، ناز و میمون.
گیاهان روز کوتاه SDP: Short Day Plants برای به گل رفتن نیاز نوری کمتر از 12 ساعت دارند. که در نقطه مقابل گیاهان روز بلند قرار میگیرند، مثل گل داودی.
گیاهان بی تفاوت به طول روز NDP: Neutral Day Plants برای نگهداری در منزل بسیار مناسب و مطلوب هستند. مثل گل حنا یا بگونیا که حساسیتی نسبت به طول روز ندارند و در تمام طول سال گل دارند.
به گل بردن گیاهان روز کوتاه در طی ساعات روشنایی، طولانی به موضوع کنترل نور بر میگردد. در کشور ما با توجه به اینکه روزهای آفتابی خیلی زیاد است باید نیاز نوری گیاه شناخته شود تا گیاه در طی روزهای بلند بخوبی از رشد رویشی برخوردار باشد. لذا گیاهان روز کوتاه را به وسیلهی پایین آوردن ساعات روشنایی، بوسیله یک پرده تیره رنگ، و اصطلاحاً ایجاد شبهای طولانی به گل میبریم.
منبع: http://www2.irib.ir/amouzesh/j/poudman_gol/taksir/page02.htm